- МЕХАНИКА РАЗВИТИЯ
- МЕХАНИКА РАЗВИТИЯ. Содержание: История......................18 Материалы и методы исследования........20 Проблема детерминации.............22 Два основных типа формообразования......26 М. р. и регенерация................30 Практическое значение М. р............31 Механика развития, область биологии, изучающая законы образования структур в индивидуальном развитии организма. Теоретически и экспериментально М. р. обоснована Вильгельмом Ру. Основной особенностью новой области В. Ру считал каузально-аналитический метод исследования морфол. проблем. Название, предложенное им, равно как и понимание задач новой области, вызвало целый ряд возражений (О. Гертвиг, Г. Вольф и др.).Прежде всего возникло неправильное представление, будто новая область должна законами механики объяснить явления развития в биологии. Ру указал напр. то, что слово «механика» употребляется им не в узком смысле физики, но в том широком, в каком Кант относит к механике каждое явление, подлежащее причинности. В программном всту- плении к первому тому основанного им Архива механики развития (Archiv fur Ent-wicklungsmechanik der Organismen) Ру пишет (1895): «Механика развития, или каузальная морфология организмов... есть учение о причинах возникновения, сохранения и обратного развития структур», и далее: «всякое подчинение причинности тех или иных явлений согласно оценке механизма, данной Кантом и Спинозой, должно быть обозначено как механическое явление. Т. о. учение о причинах становления можно назвать механикой развития». Сужение задач исследования анализом явлений развития без учета необходимости синтеза и одностороннее понимание М. р. как причинной морфологии организмов характеризует идейную-связь Ру с механистическим материализмом и с методологией Канта, поскольку последний выступал как механист. Стихийно Ру однако перерастал механистическое понимание явлений развития, и в его методологических взглядах появляются концепции материалиста-диалектика. Ряд авторов предложил для новой дисциплины другие названия, напр. физиология развития (Г. Вольф» и Г. Дриш). Жак Лёб назвал физиологич. морфологией область, по сути дела совпадающую с механикой развития В. Ру. Пользуются также индиферентными терминами г экспериментальная эмбриология, экспериментальная морфология. История. Начало экспериментальных исследований морфогенеза относится к 18 в.— знаменитые опыты Трамблея по регенерации гидры (1744), опыты Бонне, Спалланцани и др. Прочную основу эта область получила с победой эволюционной идеи во второй половине 19 в., когда за развитием описатель-но-морфол. исследований последовал второй этап—изучение процесса формообразования.. Начало было положено почти одновременно-В.Ру, начавшим в 1883 г. свои эксперимен-тально-эмбриол. работы, а в 1888 г. опубликовавшим свою историческую работу («Об искусственном получении полузародышей после разрушения одного из первых двух шаров дробления и о позднейшем развитии недостающих частей тела»), и Л. Шабри во-Франции (1887), опубликовавшим замечательное исследование по экспериментальной эмбриологии асцидий. Видную роль в развитии данной области сыграла Неаполитанская зоологическая станция, на к-рой одновременно работали ученики В. Ру— Гербст, Дриш, через посредство которых и работавших на станции американцев Е Вильсона, Т. Моргана новая область перешла в Америку, где уже независимо работал Ж. Лёб. Начало 20 в. характеризуется прежде всего усложнением и обогащением техники исследования: выработка Шпеманом метода перешнуровок, введение им микроопераций стеклянными инструментами, метод прижизненных маркировок частей зародыша В. Фохта, а в самое последнее время— сочетание метода культуры тканей с микрохирургией (лаборатория Стренджвейса в Кембридже). Вместе с тем этот период принес существенно новое и в теоретической области. В первую очередь следует отметить-учение об организаторах Г. Шпемана. В ис- тории эмбриологии большую роль сыграли представления о преформации и эпигенези-се. Ру остановился на точке зрения преформизма, но, не упуская из виду значения эпигенетического момента в развитии, он предложил говорить о двух типах образования органов—о развитии при помощи са-модиференцировки и о зависимости дифе-ренцирования. В первом случае выявляется момент преформации, во втором—эпигене-зиса. , Особую роль в истории механики развития сыграл Г. Дриш. Создав себе определенную модель материализма (крайне вульгарного), он в борьбе со своим противником скатился к признанию абсолютной автономности жизненных явлений и наличию изначальной целесообразности (энтелехия), явившись одной из самых реакционных фигур в биологии последних десятилетий. Основой теоретических взглядов Дриша является так наз. эквипотенциальная система. С фактической стороны дело сводится к тому, что на ранних стадиях нередко часть зародыша (или часть тела животного при регенерации) дает целый организм,—напр. один из бластомеров 4-клеточного стадия у морского ежа дает целую личинку. Дриш делает отсюда вывод, что яйцо изотропно, равномерно по всем направлениям, а отсюда—дальнейшее следствие, что силы, строящие зародыш,—нематериального порядка. Это положение развито им в т. наз. двух доказательствах витализма. Суть их сводится к тому, что если бы живые существа представляли трехмерные машины, то при отнятии у такой машины части, оставшаяся часть не могла бы восстановить целой структуры. Так как однако такое восстановление происходит (при опытах с регуляцией над зародышами и при регенерации взрослых организмов), то это указывает по мнению Дриша на невозможность объяснить явления в живом организме на основах физики и химии,.—Заколдованный круг, в котором находится Дриш, таков: или живые организмы—трехмерные машины, все процессы в которых сводятся без остатка к физико-химическим явлениям, или они управляются нематериальными силами. Отправляясь от такой основы, Дриш создал ряд своих ирреальных понятий. Только в самое последнее время, после всей надуманности и путаницы понятий, которыми он блистал в течение 40 лет, Дриш пришел к следующим выводам: машинная теория не представляет антипода к витализму; антиподом к витализму было бы понятие «автоматический»; машинная теория и есть виталистическая теория, т. к. машина немыслима без того, кто ею управляете живых организмах таким управляющим моментом является энтелехия. Задачи и объем М. р. Практическая задача, цель М. р.—это возможность управления явлениями возникновения признаков в процессе онтогенеза. Очевидно, что к М. р. •относится любое явление формообразования независимо от возраста (от яйцеклетки и до ■смерти организма). Но следует подчеркнуть, ■что лишь тогда мы имеем дело с М. р., когда речь идет об исследовании законов формообразования, а не о констатировании факта перехода одного законченного состояния в другое. В частности в области эндокринологии следует разделять возникновение новой структуры от выявления при помощи тех или иных раздражителей уже имеющихся структур. Учение об опухолях, относящееся в наст, время к патологии, можно думать войдет в область М. р. Из других дисциплин особенно близка к М. р. генетика, изучающая закономерность наследования признаков. Геккер предложил слить М. р. с фено-генетикой, т. е. рассматривать М. р. как область, к-рая исследует осуществление признака. Такое сужение задач нельзя признать удачным—М. р. изучает законы морфогенеза .и независимо от феногенетического осуществления признака (регенерация, бесполое размножение). Т. о. в наст, время основным ядром М. р. является область экспериментального исследования зародышевого развития, регенерации и бесполого размножения, а также в ряде случаев изучение морфогенеза во взрослом состоянии у высших позвоночных. М. р. уже в настоящее время играет огромную роль для практического врача, занимаясь причинным изучением тератогенеза. Материал и методы исследования. Излюб-леннейшим материалом для исследования зародышевого развития продолжают быть яйца земноводных, лягушки и др. бесхвостых амфибий и еще более пригодного для ранних стадиев полосатого тритона, особо ценные качества к-рого были показаны Шпеманом и позволили ему открыть новый этап в М. р. Другой ценный материал — яйца морских ежей—со времени ранних работ бр. Герт-вигов является наиболее частым и наиболее пригодным объектом для исследования М. р. беспозвоночных. Для вопросов регенерации излюбленными объектами являются гидроидные полипы (в частности пресноводная гидра) и плоские черви (планарии), из позвоночных—тритон. В последнее время начал осуществляться синтез методов культуры тканей и М. р., в связи с чем стало возможным пользоваться зародышами теплокровных птиц; в самое последнее время в лаборатории Стренджвейса начаты операции и над зародышами млекопитающих. Это открывает путь к исследованию регенерации у высших позвоночных (см. ниже). Методы исследования М. р. весьма многообразны — различные приспособления для исследования действия силы тяжести, растворы солей, центрифугирование, избирательное уничтожение определенных участков зародыша ультрафиолетовыми лучами (аппарат Чахотина) и т. д. Особенное значение приобрела микрохирургия (или так наз. микрохирургическ. техника) в исследованиях над эмбриональными трансплянтациями. Метод эмбриональных трансплянтаций был введен в 90-х гг. Борном. Борн, как и целый ряд позднейших исследователей, имел дело с поздними зародышами и пользовался металлическими инструментами—тонкими иглами, иглами, обточенными в виде ножей, тонкими ножницами и т. п. Крупной заслугой Шпемана является введение стеклянного инструментария для оперирования ранних зародышей. Важнейшие типы инструментов Шпемана видны на прилагаемом рисунке (рис. 1). Прежде всего это стеклянные иглы, служащие для разреза тканей; волосяная петля, применение к-рой в наст, время крайне разнообразно, представляет согнутый кусочек детского волоска, вставленный в тонко оттянутый конец пипетки. В зависимости от размера петель ими можно не только ориентировать зародышей, но и делать все тончайшие операции до т. наз. обмена мелких кусочков {см. ниже). Шпеман усовершенствовал также метод пере шнуровок для яйца тритона. Он пользовался тонкими волосками, делая из них петлю с морским узлом (рис. 2). Перешнуровки дают возможность исследовать время детерминации, различные вопросы каузального генеза двойных уродств (уклонения одного из близнецов, частота situs inversus viscerum etc.), границу т. наз. организационного поля, процесс гаструляции. В самое последнее время Г. А. Шмидту удалось, работая в лаборатории Шпемана, выработать метод перешнуровки яиц бесхвостых амфибий. [См. микрофотографии на отдельной табл. (рис. 1)—перешнуровка яиц бесхвостой амфибии—жерлянки (Bombina-tor pachypus): A—стадий ранних гаструл (зародыши связаны узким мостиком); В—■ поздние гаструлы; С—стадий нервных валиков; D—дорсо-вентральная перешнуровка; правый зародыш—т. наз. брюшной кусок.] Важную роль в исследовании по М. р. играет т. н. метод прижизненных маркировок. Примененный впервые американцем Гудел ом (Goodale), он был развит в недавнее время (1922—25) В. Фохтом. В существенных чертах метод состоит в том, что кусочки агар-агарапропитываются раствором какой-нибудь витальной краски (чаще всего Nilblausulfat, иногда также Neutralrot) и затем накладываются при помощи особых приспособлений на поверхность зародыша в тех или иных участках. Этот метод дает возможность устанавливать быстроту размножения различных участков зародыша, что позволило Фохту произвести тонкий анализ процесса гаструляции и нарисовать карту презумптивных зачатков в яйце хвостатых и бесхвостых амфибий. Этим методом воспользовался ряд авторов для исследования
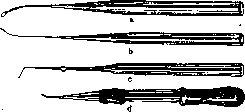
Рисунок 1. Главнейшие стеклянные инструменты
для микрохирургии ранних'зародышей: а— волосяная петля; b и с—стеклянные иглы; d—микропипетка. вопросов расположения зачатков на ранних стадиях (например зачатков глазной чаши на стадии нервной пластинки). Наконец очень видную роль в М. развития очевидно сыграет метод культуры тканей. С помощью его Уоддингтону (Waddington) удалось в 1930 г. получить индукцию нервной пластинки у зародышей курицы и утки. Этот метод дает возможность также культивировать части зародышей и доводить их до поздних стадиев. На анат. конгрессе в Амстердаме (1930) Фелл, глава лаборатории Стренджвейса, демонстрировала зачатки костей курин. зародышей, доведенные ею в питат. среде до полного сформирования и диференцировки гист.
Рисунок 2. Перешнуровка яиц тритона петлей
из детского волоска: а—до начала дробления; Ь—после снятия петли (внутри оболочки два однояйцевых близнеца). строения. Раньше для культивирования частей зародыша пользовались методами Мер-фи-Данчаковой. Изучение факторов, усиливающих и угнетающих процессы регенерации, составляет одну из ближайших практических задач современной М. развития. Проблема детерминации. Основная проблема морфогенеза в отношении к клетке поставлена была А. Вейсманом и разработана в его же учении о зародышевой плазме. Затем В. Ру перенес в механику развития Вейсма-новское представление о наследственной неравноценности ядерных делений и о соответствии делений ядер структуре образующегося организма. Первая работа по М. р. Ру «О времени определения главных осей зародыша лягушки» вышла в 1883 г., первые исследования с уколом бластомеров—в 1885 г., работа Шабри—в 1887 г.Шабри придумал ряд остроумных приборов и инструментов, к-рыми он избирательно уничтожал различные клетки дробящегося яйца и наблюдал после дующие стадии развития. Постановка проблемы у Ру была следующей: когда определяется судьба частей зародыша? Он начал с двуклеточного стадия. Предстояло решить, эквипотенциальны ли обе первых клетки дробящегося яйца или детерминированы соответственно определенным частям организма. Раньше всего он установил, что при устранении всех привходящих обстоятельств первая плоскость дробления совпадает со срединной плоскостью зародыша. К этому утверждению спустя 20 лет присоединился Браше (Brachet). Шпеман на основании исследований по перешнуровке яиц тритона пришел к выводу, что в 3/з—8/« случаев срединная плоскость совпадает со 2-й бороздой, в х/4—Vs—с 1-й бороздой. Фохт при помощи метода витальных маркировок мог отметить еще большую вариацию в отношении между 1-й бороздой и срединной плоскостью зародыша. Следует также отметить, что у ряда других животных имеется еще более редкое совпадение плоскостей дробления с плоскостями симметрии. Жюлен и ван Бенеден (Julin, van Beneden) установили у асцидий совпадение 1-й плоскости дроб- ления с плоскостью симметрии. У ктено-фор 1-я плоскость дробления совпадает с желудочной плоскостью, а 2-я—с плоскостью щупалец. В ряде случаев плоскость симметрии будущего зародыша выявляется в структуре яйца еще до оплодот- 2 ворения, что заставило Конкли-на (1929) заметить,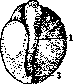
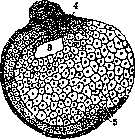
Рисунок 3. Опыты Ру: a—Hemiembryo dexter; ft—Hemiembryo sinister; 1 и г—нервный валик; 3—в а незакрытая часть бластоиора, в Ь—первичная кишка; 4—хорда; 5—недиференциро-ванные клетки. В Ъ уколотая часть полностью превратилась в клеточную массу—начало постгенерации.
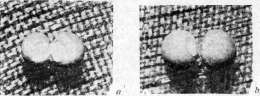
что «онтогения начинается до оплодотворения». Эксперименты Ру заключались в уничтожении раскаленной иглой одного из бла-стомеров двуклеточного стадия и изучении судьбы оставшейся в живых половины яйца. Он нашел, что как правило оставшаяся живой половина яйца дает половину зародыша. Проделывая то же с 4, 8 клетками и т. д., он нашел, что части яйца детерминированы по отношению к будущему организму (рисунок 3). С возражением выступил О. Гертвиг, показавший,что если освободить из оболочек живую половину, то она дает целого зародыша, что следовательно половинный зародыш развивается лишь в присутствии убитой половины. Т. Морган показал также, что если, не удаляя убитой половины, перевернуть яйцо, то из оставшейся половины развивается целый зародыш. Морган воспользовался мыслью эксперимента Шульце, к-рый показывает недостаточность Ру-Вейс-мановских представлений. Опыт Шульце заключался в том, что яйцо лягушки, зажатое между стеклянными пластинками, переворачивалось незадолго до наступления первой борозды деления. В таком положении оно оставалось до стадия бластулы (см.), т. е. до сравнительно позднего стадия развития. В результате в большом проценте случаев развивались двойниковые образования. В последнее время опыт Шульце был повторен Шлейпом (Schleip) на большом материале с несколько усовершенствованной методикой. Эти авторы упростили метод Шульце: яйцо лягушки зажимается между двумя предметными стеклами, между к-рыми по концам вложены полоски пластелина; толщина последних приблизительно равна диаметру яйца. Тем самым устраняется необходимость укрепления предметных стекол особыми резиновыми кольцами, как это делал Шульце. Эти опыты показали, что двойниковые образования получаются в известном проценте случаев не только при переворачивании яйца в начале дробления, но и в продолжение ранних стадиев до восьми клеток включительно. Словом ряд фактов показывал, что у лягушки говорить о соответствии между делениями ядра и структурой зародыша невозможно. В. Ру наблюдал, что во многих случаях полузародыши превращались в целых, и назвал этот процесс дополнительным развитием, или постгенерацией. Он вывел отсюда свои чрезвычайно важные понятия—самостоятельного диференцирования, или само-диференцирования, и зависимого диферен-цирования. Первое является, по В. Ру, типичным случаем, второе—атипичным, вызванным особыми условиями эксперимента. Почти одновременно с опытами Шульце итальянскому ученому Герличка (Herli-tzka) удалось перешнуровать тонкой шелковой нитью яйцо тритона и показать, что из обеих половин развились целые зародыши. Методу перешнуровки пришлось сыграть видную роль в исследовании Ру-Вейсма-новской гипотезы наследственно неравноценных делений ядра оплодотворенного яйца. Предварительно следует указать на другой эксперимент, исторически произведенный значительно раньше, к-рый также имел решающее значение. Дело идет об опыте Жака Лёба: яйца морского ежа вскоре после оплодотворения были перенесены в гипотонический раствор (морская вода+дестил. вода 1:1). Произошло резкое набухание и разрыв оболочки. Часть содержимого вышла в воду, приняв круглую форму (так на-зыв. экстраоват) (рисунок 4). В некоторых случаях ядро первоначально находилось в-одной половине яйца (внутри оболочки) и здесь проделывало известное количество делений. Затем на известном стадии одно из дочерн. ядер переходило в экстраоват, который таким образом получал продукт 2-го, 3-го или 4-го делений, т. е. Vs. г11в, 1/32,часть ядра оплодотворенного яйца. Но тем не менее из экстраовата развивался нормальный зародыш. В недавнее время проверка Ру-Вейсмановского правила была сделана Шпеманом (рисунок 5). Из его опытов следовало, что первая борозда разделила яйцо на спинную и брюшную части. Эти исследования окончательно-показали, что детерминация частей зародыша имеет другую основу, чем это предполагал В. Ру. В частности в механизме сформирования зародыша имеет важное значение участок плазмы, так называемое с е-р о е поле, заметный у земноводных уже до начала дробления. Шпеман показал, что только часть яйца, получившая серое поле, может развить нервн. систему, осевой скелет и мускулатуру; часть, не имеющая его, представляет т. н. брюшной кусок. Важная роль плазмы в процессе морфогенеза констатирована для ряда животных. Известнейшим примером является асцидия
Рисунок 4. Схематическое изображение опыта Жака Лёба: о—яйцо до помещения в гипотонический раствор; Ь—яйцо в гипотоническом растворе — оболочка разорвалась, часть содержимого яйца вышла наружу.


1Н 4&Ф?**ъ

>%£ л; - ' ,» • / * -* У
■К? Л' • • * • .&• & и •.*- * .. • » » •» »Г w \>€fc * * • *У 4 • zffi- 'ил , • л • *• *%*•
Рисунок 1 —3. Механика развитии (объяснение—в тексте). Рисунок 4. Спорадический кретинизм (10 л.): « — до лечения; b — через 2 недели после лечения тиреоидином; с через 4 мес. после лечения тиреоидином. Рисунок 5. Спорадический кретинизм (21 г.): ан b — до и после лечения тиреоидином. Рисунок 6. Микседема: а и b — до и после лечения. Рисунок 7. Атрофия и сморщивание (а) одной извилины головного мозга при детском параличе.
К ст. Механика развития, Микрогирпя, Микседема, Stvela partita, у которой Конклин в яйце до* начала дробления различает 5 различно окрашенных плазм [см. отд. табл. рис. 2— яйцо асцидии Styela (Cynthia) partita: A— сбоку; В—сзади; С—двуклеточный стадий сбоку; 1—светлая плазма (мезенхима); 2— желтый полумесяц (мезодерма); 3—-направительные тельца; 4—материал для хорды и нервной трубки]. Эти плазмы попадают -затем в части зародыша, соответствующие различным органам. Этот случай интересен также и в том отношении, что окончательное распределение плазм связано с процессом созревания яйца. Ф. Лилли (Lil-lie) удалось воздействием определенных со-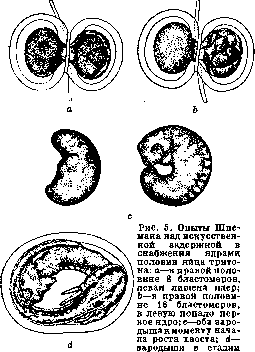
личинок с развитыми жабрами. Личинки доведены до 140 дней, и к моменту фиксации никакой разницы между ними нельзя было заметить, между тем левая развилась из половины оплодотворенного яйца, имевшей Vie ядра, а правая—i5/i6.
лей вызвать в яйце многощетинкового червя Chaetopterus диференцировку без дробления, получить одноклеточную бластулу, гаструлу и трохофору, т. е." образование ресничного покрова, соответствующего нормальному покрову. Петерфи (Peterfi) удалось вынуть при помощи микроманипулятора ядро из яйца аксолотля и получить все же развитие до позднего стадия дробления. Вильсон (Wilson) в своем известном опыте над яйцами моллюска Dentalium показал определенное значение плазматической, так наз. полярной лопасти в детерминации теменного органа и султана (рисунок 6). Фи-шель (Fischel) показал детерминирующее значение плазмы в развитии ктенофор. В недавнее время органообразующая плазма яйца ктенофор была исследована Шпеком (Speck) при помощи метода темного поля, давшего возможность показать необычайно демонстративно ее перемещения в начале дробления. Органообразующие вещества известны для целого ряда червей. Эти факты показывают, что плазма играет важную роль в механизме развития зародыша—в процессе детерминации различных органов его. Факт важной роли плазматического компонента клетки в процессе детерминации не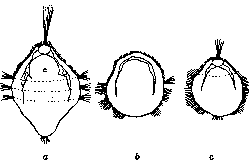
Рисунок 6. Опыты Вильсона над Dentalium: a— нормальная трохофора; 6—личинка после удаления первичной желточной лопасти; с— личинка после удаления вторичной желточной лопасти; в стадии Ъ султан совсем не развился.
стоит в противоречии с хромосомной теорией наследственности, т. к., дело идет о совершенно другой категории явлений—не о механизме передачи наследственных признаков, а о механизме морфогенеза—развития признака. Два основных типа формообразования и проблема их соотношения. Учение об организаторах, понятия лабильной детерминации и двойного обеспечения. Опыты по перешнуровке яиц тритона были начаты Шпеманом с 1898 г. и продолжались до 1904 г. В дальнейшем Шпеман занялся вопросом о детерминации глаза (рисунок 7). Как известно, в процессе развития нервной системы глазные пузыри, представляющие выросты промежуточного мозга, дорастают до эпителия, и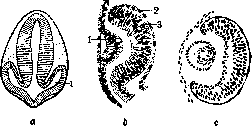
Рисунок 7. Развитие глаза у позвоночных: Fa-стадий глазных пузырей (2); Ь—ямковидный зачаток хрусталика (1) (2 и 3—два слоя возникшей глазной чаши); с—более поздний стадий—зачаток хрусталика обособился от связи с эктодермой.
в месте соприкосновения образуется ямко-образное вдавление эпителия—зачаток хрусталика. Вместе с тем наружная и дисталь-ная стенки глазного пузыря начинают вдавливаться—глазной рузырь превращается в чашу. Можно было думать, что влиянием образующегося хрусталика следует объяснить переход глазного пузыря в глазную чашу. Оказалось же как-раз обратное—у ряда амфибий впячивания хрусталика не происходит, если вырезать зачаток глазного пузыря (вырезается на стадии нервной пластинки участок, соответствующий глазному пузырю). У некоторых амфибий (съедобная лягушка) хрусталик все же развивается. В других случаях глазной бокал переносился под кожу другого места головы или туловища. У древесницы (Ну la arborea) в любом месте тела образуется хрусталик. У серой лягушки хрусталик образуется в любом месте головы. У съедобной лягушки не каждый участок кожи головы способен превращаться в хрусталик, но вместе с тем глазная чаша имеет способность индуцировать хрусталик, как показывает опыт Филатова, заменявшего у съедобной лягушки кожу над глазным пузырем участком кожи жабы, взятым из любого места головы,—происходило образование хрусталика. Эти опыты, произведенные рядом авторов (Шпеман, Льюис, Экман, Филатов), приводят к представлению, что развитие одного органа происходит под воздействием другого органа, что следовательно принцип зависимого диференцирования В. Ру может пониматься в смысле непосредственной связи, в смысле построения одного органа другим. Шпеману удалось еще в работах по детерминации глаза сделать наблюдение в новом направлении. Занимаясь обменом кусков над областью глазной чаши при помощи особого инструмента, т. наз. микропипетки, Шпеман пробовал проделывать такой обмен между кусками разных областей эктодермы у двух зародышей. Он установил, что на стадии гаструлы стойкой детерминации нет, что куски из области презумптивной нервной пластинки и из области кожи брюха после обмена продолжают развитие соответственно месту (рис. 8). Далее Мангольд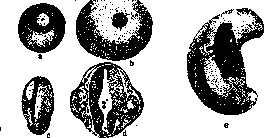
Рисунок 8. Опыты Шпемана по обмену кусками презумптивной нервной пластинки и брюшного эпителия между зародышами Tr. taeni-atus (полосатый тритон) и Тг. cristatus (гребнистый тритон): а—зародыш Тг. taeniatus с пересаженным (белым) куском презумптивного брюшного эпителия Тг. cristatus; b—то же в стадии нейрулы; d—разрез через того же зародыша на стадии первичных глазных пузырей; с—зародыш Тг. cristatus вскоре после операции с куском презумптивной нервной пластинки; е—то же на стадии первичных глазных пузырей и хвостовой почки—темный имплянтат превратился в эпидермис тела.
(Mangold) показал отсутствие стойкой детерминации на этих стадиях и по отношению к различным зародышевым листкам: кусок эктодермы, пересаженный в мезодерму, развивается соответственно месту. Шотте (Schot- te) показал то же и по отношению к мезодерме — ее способность превращаться в эктодерму. Уже в самом начале своей работы Шпеман сделал наблюдение, что иногда куски оказывают влияние на окружающие их ткани. Оказалось, что куски эктодермы„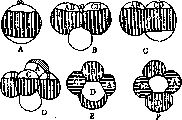
Рисунок 9. А—две гаструлы полосатого тритона, соединенные таким образом, что их срединные плоскости дивергируют кпереди; В—личинка, возникшая путем срастания этих половинок—duplicitas anterior (переднее удвоение); С—переднее удвоение у теленка; D—два зародыша полосатого тритона соединены таким образом, что их срединные плоскости конвергируют кпереди; Е—возникшая из двух зародышей (D) личинка с двумя задними концами — duplicitas posterior (заднее удвоение); F—duplicitas posterior у теленка.
взятые у дорсальной губы бластопора, пересаженные в область брюшной эктодермы» индуцируют в ней нервную пластинку. Связав этот факт с опытами по перешнуровке, говорившими за наличие в области серого-поля какой-то особой зоны, обусловливающей построение осевого комплекса, Шпеман пришел к представлению о том, что в области дорсальной губы бластопора находится так наз. организационное поле. Кусок его, пересаженный в индиферентнун> область, вызывает в ней образование высо-кодиференцированных органов. Еще две важные серии опытов укрепили его в представлении об организационном поле: им был проделан ряд опытов по сращиванию половинок гаструл. Результаты опытов по сращиванию разноименных половин, представлены на рисунке 9. Если сращивались одноименные половинки гаструл, то-материал отрезанной половинки дорсальной губы на каждой стороне дополняется обычной эктодермой. В результате возникает-зародыш с двумя нервными пластинками и двумя осевыми комплексами. Вторая серия заключалась в том, что если крыша гаструлы срезывалась и переворачивалась на. 90° или 180°, то никакого изменения в положении нервной трубки не происходило. Последующие работы Шпемана и его учеников выяснили ряд других интересных явлений—область организационного поля переходит в крышу первичной кишки, и кусок крыши первичной кишки индуцирует нервную пластинку (Marx) [см. отдельную таблицу, рис. 3: А я В—микрофотографии зародыша с индуцированной вторичной нервной пластинкой (А—ее оку; В—с вентральной стороны, на которой и находится вторичная пластинка); С—разрез того же зародыша; 1—медулярная пластинка; на обеих сторонах видны осевые комплексы—нервная пластинка, хорда, мезодерма]. Индиферент-ная ткань, помещенная в организационное поле, приобретает индуцирующие способ-- ности. Шпеман обнаружил, что кусок организационного поля вызывает в эктодерме явления, которые подобны начальным стадиям нормальной индукции. Бауцман (Bautz-mann) выяснил область распространения организационного поля и показал, что хорда также способна индуцировать нервную пластинку в эктодерме. Мангольд и Шпеман открыли замечательное явление равноценной индукции—кусок нервной трубки старшего зародыша образует нервную пластинку в эктодерме младшего. Гамбургер (Hamburger), изучая диферен-циащда нервной системы конечности, установил 4 стадия: 1) врастание нервов в почку конечности (притяжение их почкой), 2) ди-ференциацию сплетений, 3) диференциров-ку крупных нервных стволов под влиянием кровеносных сосудов, 4) диференцировку концевых веточек под воздействием мышц или кожи. Из всех этих фактов Шпеман вывел два чрезвычайно важных теоретических принципа: 1) принцип лябильной детерминации—на стадии гаструлы вся эктодерма лябильно детерминирована, что показывается опытами с обменом участков; 2) детерминация происходит по принципу двойного или множественного обеспечения; иными словами орган детерминируется по меньшей мере из двух источников, напр. нервная пластинка детерминируется из серого поля и из крыши первичной кишки. Исследования над структурой организационного центра дали лишь самые общие указания о его лятеральном и передне-заднем расчленении. Основным, но наиболее темным является вопрос о природе индукционного действия. В последнее время делались попытки выяснить свойства организационных центров. Так, Аникин (1927) показал, что губы бла-стопора являются источником митогенети-ческих лучей (см.). Этому же автору удалось констатировать излучение медулярной борозды, прекращающееся после замыкания последней. Бляхер и Замараев констатировали (1930) наличие митогенетического излучения в гипостоме и зоне почкования у гидры. Эти участки тела гидры обладают свойствами организационных центров (см. ниже). Приведенные данные не объясняют исчерпывающе механизма формообразующего действия организационных центров. Во всяком случае они могут служить материалом для суждения об этом механизме с точки зрения причин клеточного размножения в сфере действия организационного поля. Т.о. работы Шпемана и его школы показали наличие двух принципов в детерминации— это как бы два типа В. Ру (принцип самоди-ференцирования и зависимого диференциро-вания), только примененные с гораздо большей ясностью и последовательностью. В развитии любого органа существует элемент мозаики—элемент предопределения и элемент возможности изменения, т. е. регуляции. Исследования Фохта при помощи метода дефектов показали, что оба момента могут присутствовать одновременно. Проблема соотношения мозаичного развития и регуляции—или лябильной детерминации п воздействия организатора того или иного порядка—очередная проблема М. развития. Механика развития и регенерация. Исследования В. Исаева показали существование зон высокой морфогенетической активности во взрослом организме. У гидры, объекта его исследований, такой зоной является гипостом, область, от которой отходят щупальцы, окружающие ротовое отверстие, зона почкования и подошва. Геч (Gotsch) подтвердил наблюдения Исаева над гидрой. Он показал существование зон высок, активности у планарий при регенерации и бесполом размножении. Эта зона у них находится в головном отделе. Последнее наблюдение хорошо согласуется с работами американского эмбриолога Чайлда (Child), пришедшего к представлению о морфогенетической неравноценности взрослого организма другим путем. Исследуя скорость метаболизма, Чайлд нашел, что она различна в различных тканях. У целого ряда животных область усиленного обмена находится в переднем конце тела. У двусторонне симметричных животных прогрессивное падение интенсивности процессов обмена совпадает с осью животного. Интенсивность окислительных процессов определялась Чайлдом марганцовокислым калием, редуцирующимся в протоплазме с образованием коричневой окраски двуокисного соединения марганца. Процессы отравления и отмирания также совпадают с осью тела животного. Изменяется по оси и электрический потенциал. Ультрафиолетовые лучи убивают раньше всего ткани в области зон высокой активности. Это показали опыты с гидрой, у к-рой зона высшей активности, как указано выше, находится в гипостоме. Зоны высокой физиол. активности установлены у ряда животных. Помимо главной оси существуют вторичные оси, направленные к сторонам тела. Морфо-генетическое различие разных областей тела проявляется в процессе регенерации и бесполого размножения. Если из тела плоского червя вырезать кусок так, что длинная ось отрезка совпадает с длинной осью тела, тогда головной конец регенерирует по направлению этой оси. Если кусок вырезан так, что верхушкой касается длинной оси, тогда эта верхушка является местом наибольшей физиол. активности. При регенерации головы должна быть известная разница в скорости реакции между передней отрезанной поверхностью и концом куска. Все эти факты привели Чайлда к так наз. теории осевых градиентов, согласно которой интенсивность морфол. и физиол огич. процессов падает по оси тела животного; у исследованных им объектов—в направлении спереди назад. Выводы, которые делает Чайлд, таковы: зрелый организм Не представляет предопределенной гармонии независимых частей. Между различными органами существуют взаимн. влияния. Основой целостности организма является физиол. доминант а—область, господствующая в системе организма. Физиол. доминирование варьирует в своем направлении; при этом может происходить изоляция частей организма, к-рые становятся при известных условиях новыми индивидуумами. В последнее время Чайлд намечает параллели между теорией осевых градиентов и учением об организационных центрах в развитии земноводных. Один из учеников Чайл-да показал наличие зоны высокой физиол. активности в области организационного центра. Чайлд считает, что организационный центр в основе не отличается от других доминирующих областей тела. На анат. конгрессе в Амстердаме (1930) Фохт сделал сообщение о соотношении регуляции и регенерации и показал, что регенерация — специфическое свойство, развивающееся лишь на поздних стадиях. Для высших позвоночных проблема возникновения регенерации или решение вопроса о том, почему регене-рационная способность не осуществляется,— одна из величайших задач современной М. р. В плоскости исследования этих специфических закономерностей процесса регенерации особый интерес представляют работы школы Гийено. Шотте показал, что процессы регенерации у тритона зависят от нервной системы — регенерации не наступает, если лишить конечности иннервации и именно, как это выяснено им в ряде интересных экспериментов, в отсутствие симпатической иннервации. Конечности, неподвижные и лишенные чувствительности (с перерезанными двигательными и чувствительными нервами), но сохранившие симпатиче-•скую иннервацию, регенерируют; конечности же, сохранившие чувствительность и функцию движения, но лишенные симпатической иннервации, не регенерируют. Другие серии опытов установили, что симпатический нерв не имеет самостоятельного значения; этот нерв лишь передает импульсы, идущие от желез внутренней секреции. У взрослых тритонов и личинок саламандр после разрушения гипофиза конечности не регенерируют при неизмененной иннервации. Это направление работ не касается факторов диференциации. Исследование последних в указанной школе привело к представлению о т. н. территориях. В регенерации конечности такой территорией являются мягкие ткани, окружающие скелет. При удалении их регенерации не наступает. Работы же Шотте о влиянии на процесс регенерации желез внутренней секреции и симпатического нерва говорят о центральном компоненте процесса. В данном отношении это одно из первых указаний на роль эндокринных желез в процессах морфогенеза при регенерации. Практическое значение М. p. I. M. р. и тератология. Работы Шпемана оказали большое влияние на исследование тератогенеза. В одном из своих первых исследований Шпеман описывает случай циклопического уродства и анализирует его. Ге-нез циклопии явился предметом ряда исследований других авторов. Целый ряд исследований, вышедших как из лаборатории Шпемана, так и из других ин-тов, посвящен различным вопросам генеза двойниковых уродств. При помощи метода перешнуровок Шпеману и Фалькенбергу удалось подойти к такому тонкому вопросу, как происхождение явлений асимметрии. Основные типы двойных уродств (переднее, заднее и крестообразное)" получают прекрасное объяснение в связи с принципом организационного по- ля—дивергирование осей гаструлы дает переднее, конвергирование — заднее, направление друг на друга — крестообразное двойное уродство (рисунок 9). Детальнее о работах по М. р. в связи с тератогенезом—см. Тератология. II. М. р., хирургия и патология. Отношение 31. р. к хирургии вытекает из всего сказанного по поводу регенерации. Метод культуры зародыша в питат. среде открывает здесь новую эру. Вопросы регенерации и условий, в к-рых она протекает, подлежат дальнейшему изучению. Хирургия будущего должна пользоваться данными М. р. для овладения процессами морфогенеза.—В области патологии онкология можно думать получит освещение из данных М. р., когда будут расшифрованы и поняты такие проблемы, как явления целостности, соотношение частей в организме, сущность процесса диференциации, явления физиол. изоляции. Все эти проблемы ждут исследования и решения и составят основу для познания опухолевого процесса. Пока сделаны только первые шаги на пути к исследованию этих труднейших проблем. Лит.: Вайсман А., Лекции по эволюционной теории, М., 1918; Конклин Е., Наследственность и среда, М., 1928; D г i e s с h H., Die Phy-siologie der tierischen Form, Erg. d. Physiologie, B. V, 1906; Dtirken В., Grundriss der Entwicklungsme-chanik, В., 1929; Haeckejr V., Entwicklungsge-schichtliche Eigenschaftsanalyse (Phanogenetik), Jena, 1918; Loeb J., Untersuchungen zur physiologischen Morphologie der Tiere, Wurzburg, 1891—92; M a n-go 1 d Q., Das Determinatioiisproblem, Erg. d. Biol., B. Ill, 1928; Morgan Т., Experimental embryology, New York, 1927; Roux W., Einleitung, Arch. f. Entwicklungsmechanik, B. I, p. 1—38, 1894; он же, Fur unser Programm u. seine Verwirkli-chung, ibid., B. V, p. 1—80 u. 219—342, 1897; о н ж е, Die Entwicklungsmechanik, ein neuer Zweig der biologischen Wissenschaft, Vortrage u. Aufsatze iiber Entwicklungsmechanik, Heft i, Lpz., 1904; он же, Terminologie der Entwicklungsmechanik der Tiere u. Pflanzen, Lpz., 1912; Weiss P., Entwicklungs-physiologie der Tiere, Dresden—Lpz., 1931; Wilson E., The cell in development a. heredity, chapt. XII—XIV, N. Y., 1928 (лит.). Серия монографий.— Roux 'Vortrage u. Aufsatze iiber Entwicklungsmechanik, Lpz., с 1904 (продолжение под назв.—Abhandlungen zur Theorie der organischen Entwicklung, В., с 1926). Периодическое издание.—Roux' Archiv f. Entwicklungsmechanik der Organismen, В., с 1895. См. также лит. к ст. Клетка и Эмбриология. Г. Шмидт.
Большая медицинская энциклопедия. 1970.
МЕХАНИКА РАЗВИТИЯ — раздел эмбриологии, изучающий при помощи опытов на живых зародышах причинные механизмы индивидуального развития организмов. Основанная В. Ру в 80 х гг. 19 в. М. р. бурно развивалась в 1 й трети 20 в. Начиная с 40 х гг. в результате сближения М. р … Биологический энциклопедический словарь
Механика развития — раздел биологии, изучающий причинные механизмы индивидуального развития организмов. Основанная в 80 х гг. 19 в. немецким учёным В. Ру М. р. бурно развивалась в 1 й трети 20 в. Начиная с 40 х гг. в результате сближения М. р., цитологии,… … Большая советская энциклопедия
МЕХАНИКА — (от греч. mechanike (techne) наука о машинах, искусство построения машин), наука о механич. движении матер. тел и происходящих при этом вз ствиях между ними. Под механич. движением понимают изменение с течением времени взаимного положения тел или … Физическая энциклопедия
Механика грунтов — строительная дисциплина, изучающая грунты основания и их взаимодействие с сооружениями. Механика грунтов, вместе с инженерной геологией и охраной природной среды составляет особый цикл строительных дисциплин, работающих с материалами природного… … Википедия
Механика грунтов — научная дисциплина, изучающая напряжения, деформации, условия прочности и устойчивости грунтов, изменение их состояния и свойств под влиянием внешних механических воздействий. Геологический словарь: в 2 х томах. М.: Недра. Под редакцией К. Н.… … Геологическая энциклопедия
Механика сплошных сред — изучает движение и равновесие газов, жидкостей и деформируемых твёрдых тел. Моделью реальных тел в М. с. с. является сплошная среда (СС); в такой среде все характеристики вещества являются непрерывными функциями пространственных координат и… … Энциклопедия техники
Механика — [от греч. mechanike (téchne) наука о машинах, искусство построения машин], наука о механическом движении материальных тел и происходящих при этом взаимодействиях между телами. Под механическим движением понимают изменение с течением… … Большая советская энциклопедия
Механика — (греч. μηχανική искусство построения машин) область физики, изучающая движение материальных тел и взаимодействие между ними. Движением в механике называют изменение во времени взаимного положения тел или их частей в пространстве[1].… … Википедия
Механика — наука о движении. Изучая движение, механика необходимо должна изучать и причины, производящие и изменяющие движения, называемые силами; силы же могут и уравновешивать друг друга, и равновесие может быть рассматриваемо как частный случай движения … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
механика сплошных сред — механика сплошных сред — изучает движение и равновесие газов, жидкостей и деформируемых твёрдых тел. Моделью реальных тел в М. с. с. является сплошная среда (СС); в такой среде все характеристики вещества являются непрерывными функциями… … Энциклопедия «Авиация»
МЕХАНИКА РАЗВИТИЯ
18+
© Академик, 2000-2025
- Обратная связь: Техподдержка, Реклама на сайте
Экспорт словарей на сайты, сделанные на PHP, Joomla, Drupal, WordPress, MODx.